
Gestational Iron Supplementation Improves Fetal Outcomes in a Rat Model of Prenatal Alcohol Exposure

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
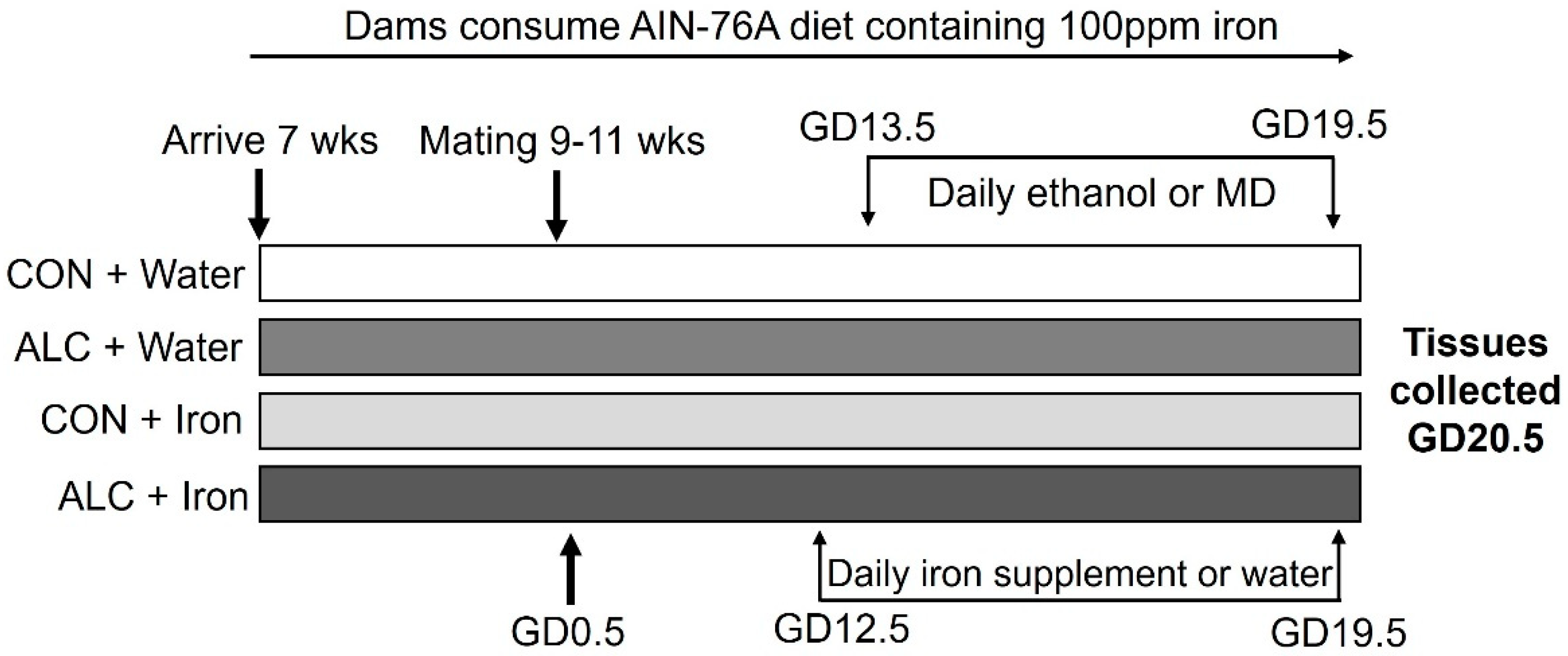
2.1. Animals and Diet
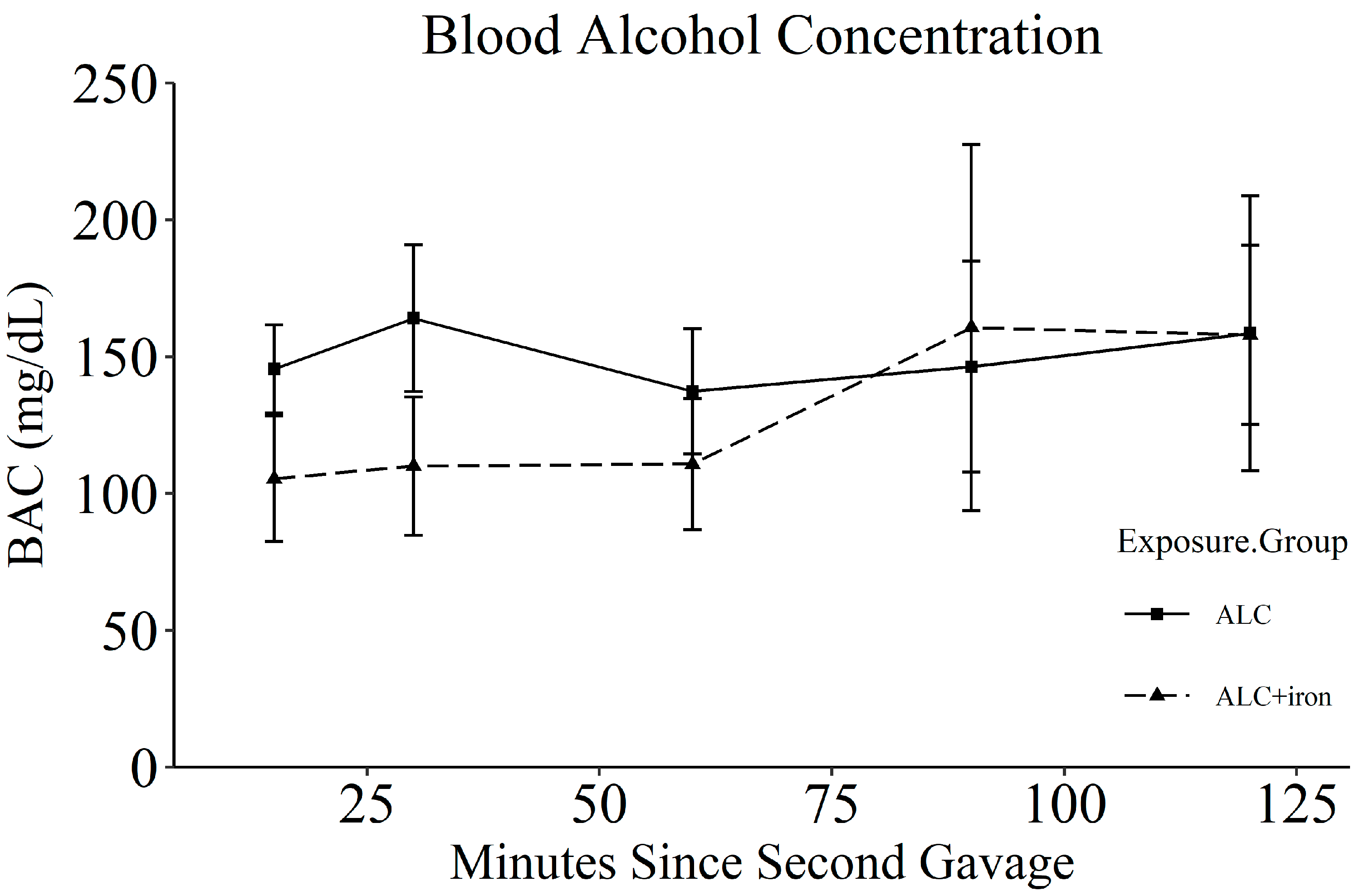
2.2. Alcohol Exposure
2.3. Iron Supplementation
2.4. Tissue Collection
2.5. Hematology
2.6. Fetal Sex Genotyping
2.7. Mineral Analysis
2.8. qPCR for Hepcidin
2.9. Thiobarbituric Acid Reactive Substances Assay
2.10. Statistical Analysis
3. Results
3.1. Maternal Gestational Characteristics
3.2. Litter and Fetal Parameters
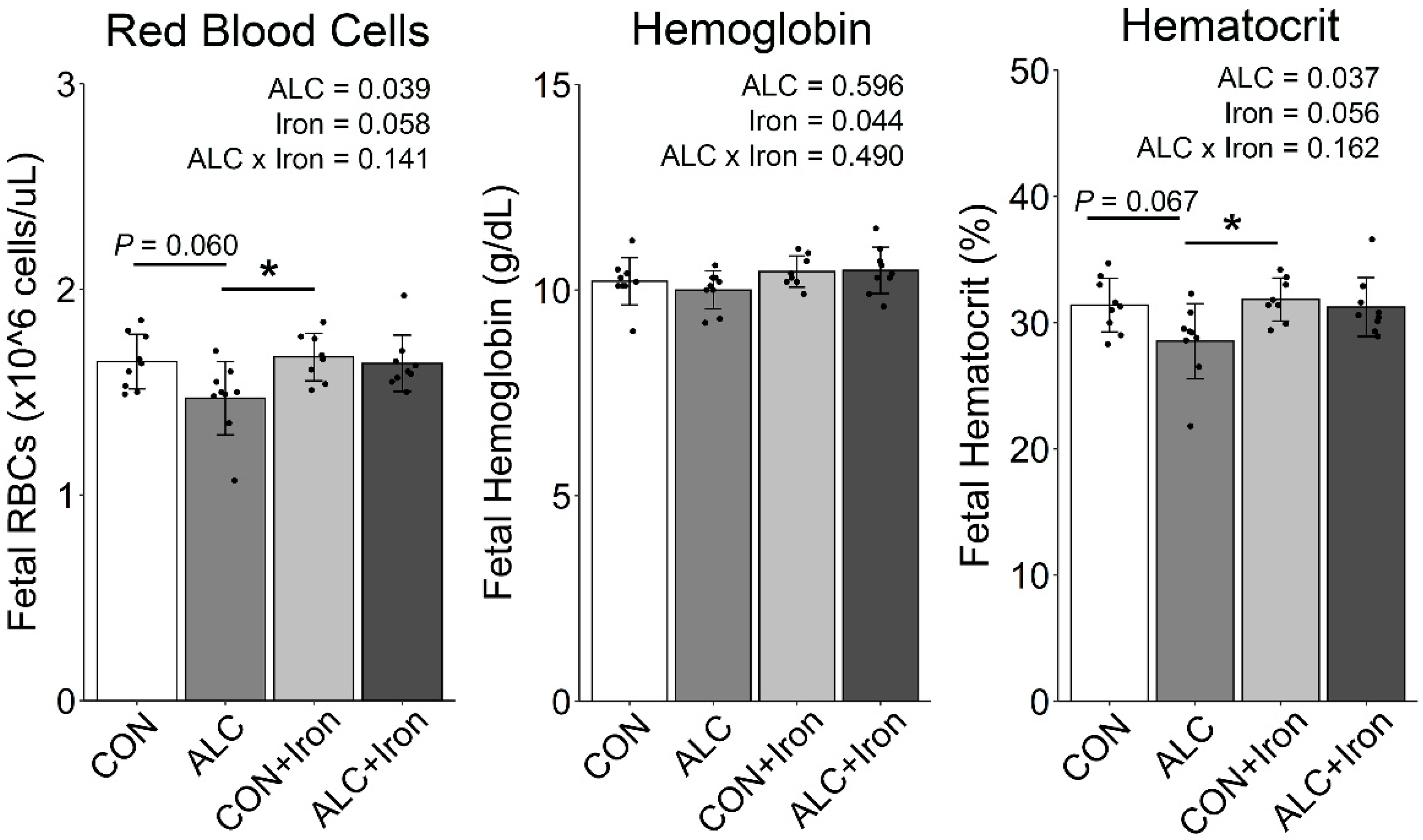
3.3. Maternal and Fetal Hematology
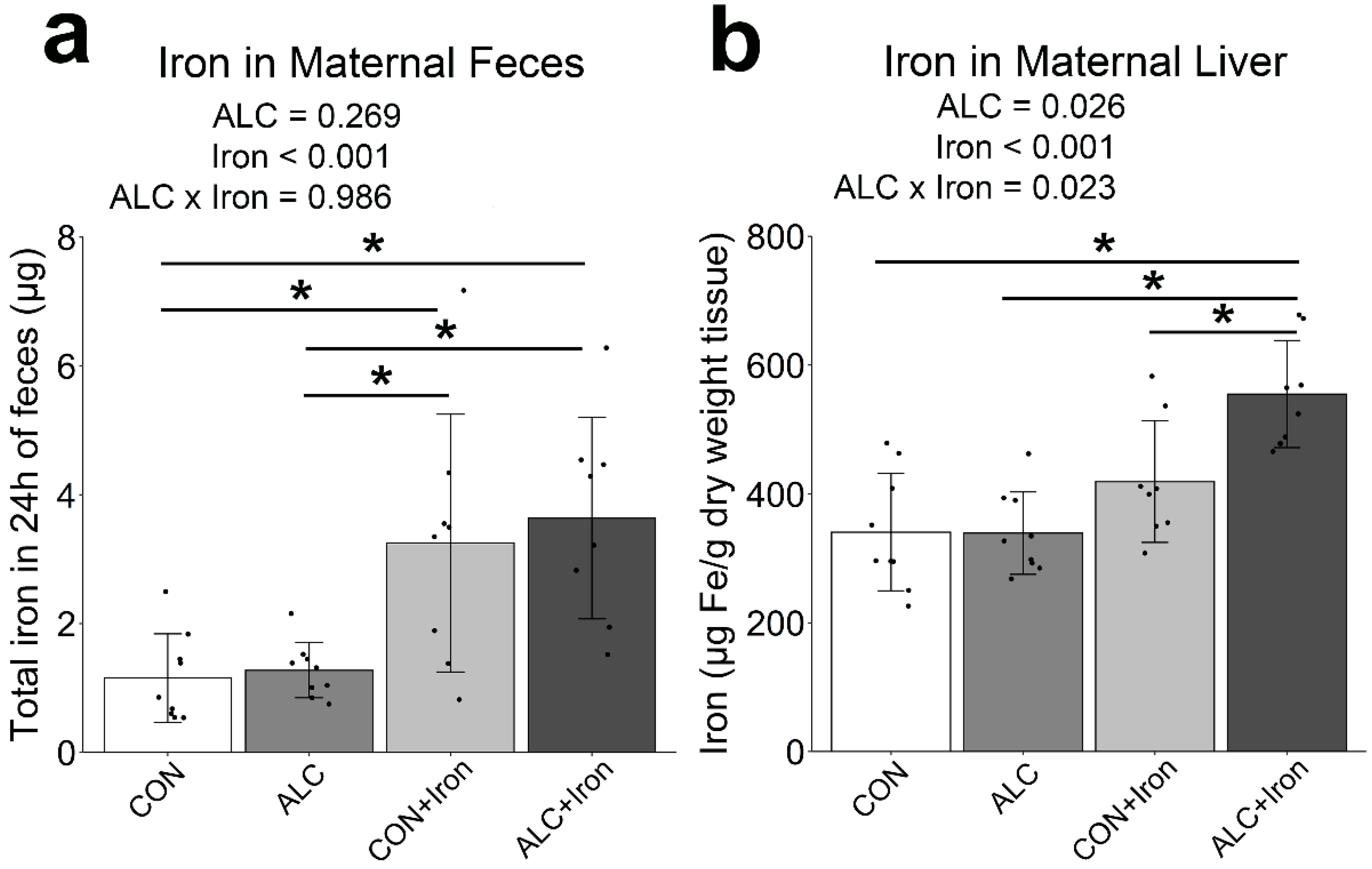
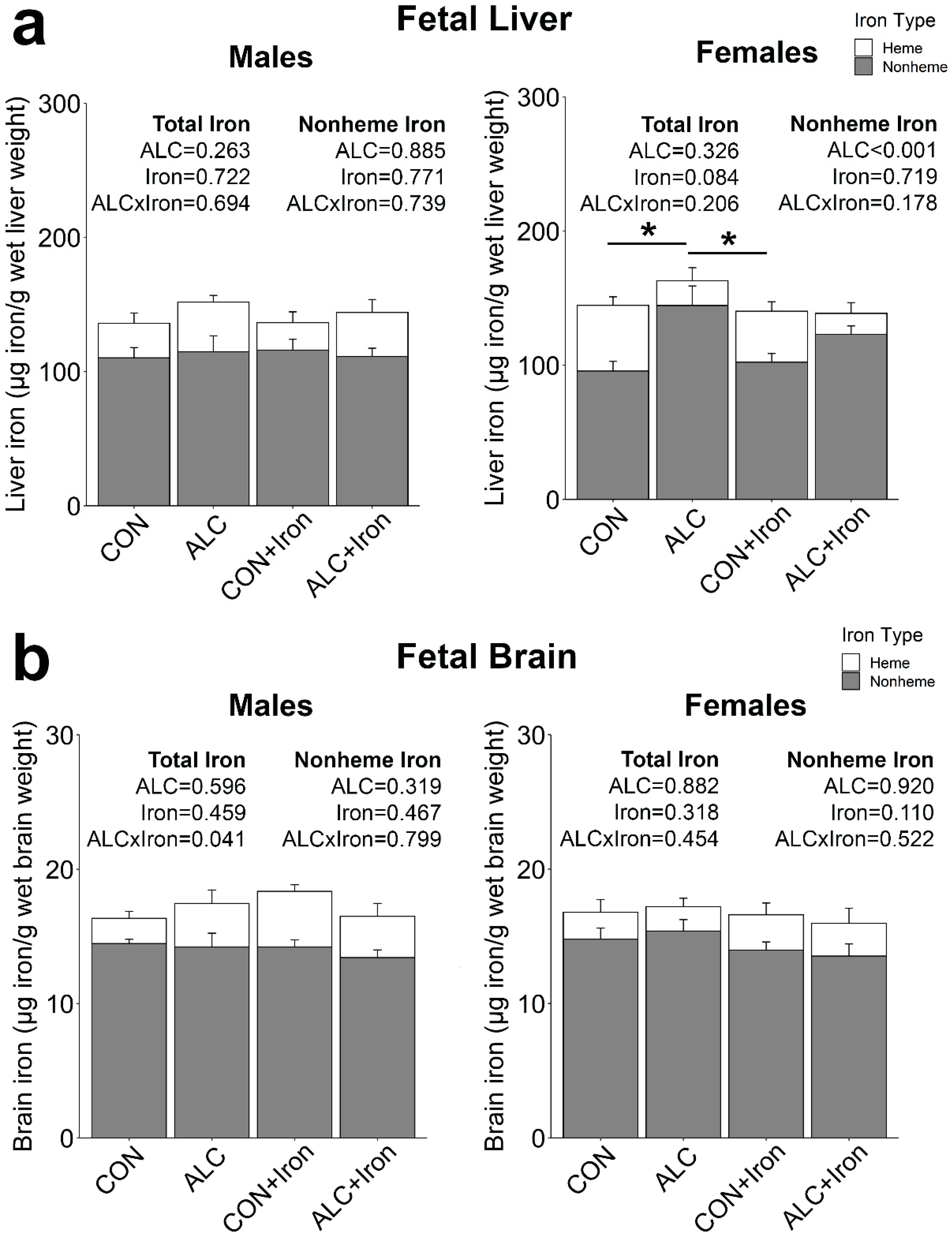
3.4. Maternal and Fetal Iron Analysis
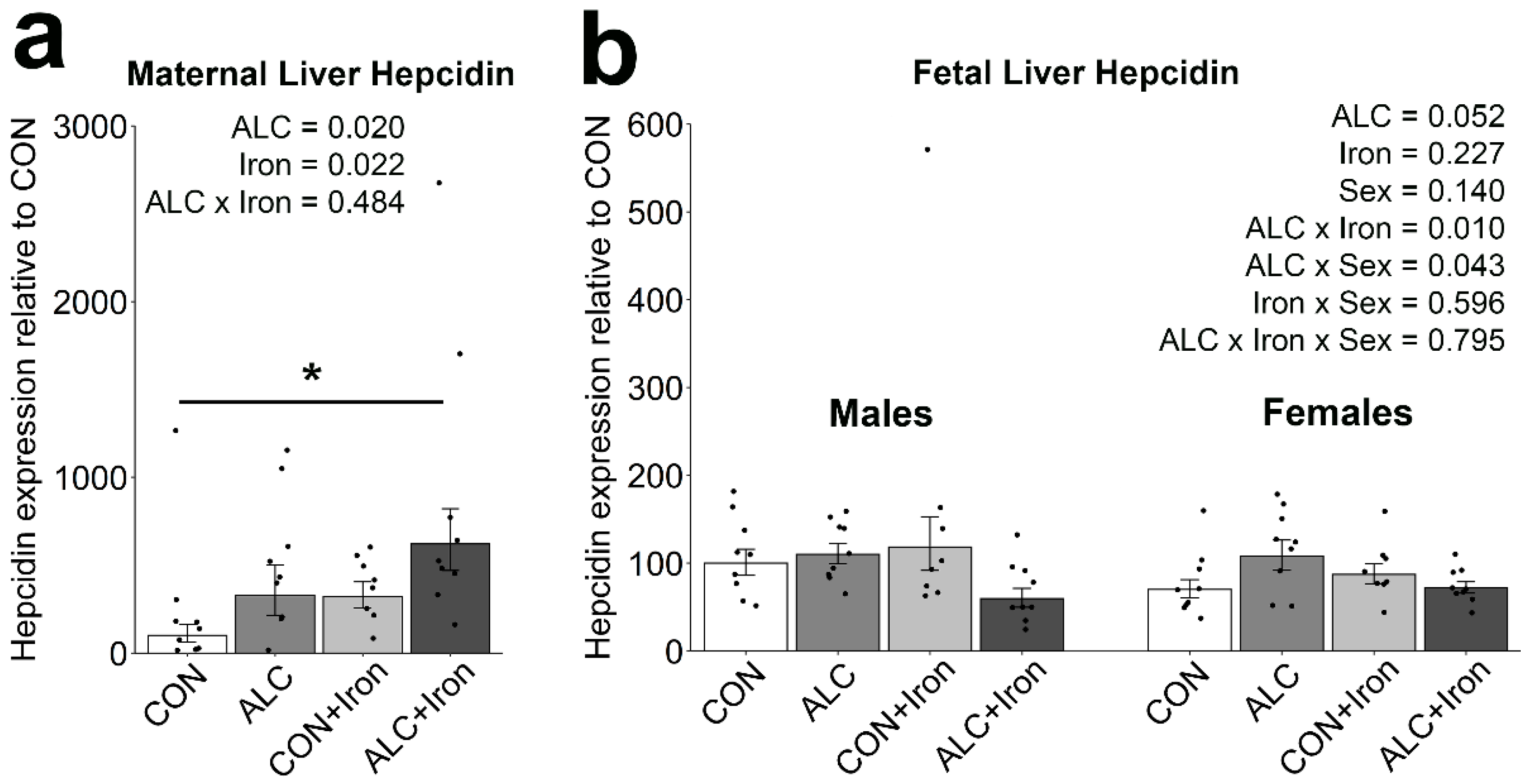
3.5. Maternal and Fetal Hepcidin
3.6. Maternal and Fetal Malondialdehyde
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hoyme, H.E.; Kalberg, W.O.; Elliott, A.J.; Blankenship, J.; Buckley, D.; Marais, A.-S.; Manning, M.A.; Robinson, L.K.; Adam, M.P.; Abdul-Rahman, O.; et al. Updated Clinical Guidelines for Diagnosing Fetal Alcohol Spectrum Disorders. Pediatrics 2016, 138, e20154256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denny, C.H.; Acero, C.S.; Naimi, T.S.; Kim, S.Y. Consumption of Alcohol Beverages and Binge Drinking Among Pregnant Women Aged 18–44 Years—United States, 2015–2017. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Greenmyer, J.R.; Klug, M.G.; Kambeitz, C.; Popova, S.; Burd, L. A Multicountry Updated Assessment of the Economic Impact of Fetal Alcohol Spectrum Disorder: Costs for Children and Adults. J. Addict. Med. 2018, 12, 466–473. [Google Scholar] [CrossRef]
- Denny, C.H.; Acero, C.S.; Terplan, M.; Kim, S.Y. Trends in Alcohol Use Among Pregnant Women in the U.S., 2011–2018. Am. J. Prev. Med. 2020, 59, 768–769. [Google Scholar] [CrossRef] [PubMed]
- Young, J.K.; Giesbrecht, H.E.; Eskin, M.N.; Aliani, M.; Suh, M. Nutrition implications for fetal alcohol spectrum disorder. Adv. Nutr. Int. Rev. J. 2014, 5, 675–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieff, M.K. Iron deficiency in pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 516–524. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.O.; Ru, Y. Iron status of North American pregnant women: An update on longitudinal data and gaps in knowledge from the United States and Canada. Am. J. Clin. Nutr. 2017, 106, 1647S–1654S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, R.C.; Jacobson, S.W.; Molteno, C.D.; Jacobson, J.L. Fetal Alcohol Exposure, Iron-Deficiency Anemia, and Infant Growth. Pediatrics 2007, 120, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.C.; Jacobson, J.L.; Molteno, C.D.; Jiang, H.; Meintjes, E.M.; Jacobson, S.W.; Duggan, C. Effects of Heavy Prenatal Alcohol Exposure and Iron Deficiency Anemia on Child Growth and Body Composition through Age 9 Years. Alcohol. Clin. Exp. Res. 2012, 36, 1973–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufer, E.S.; Tran, T.D.; Attridge, M.M.; Andrzejewski, M.; Flentke, G.R.; Smith, S.M. Adequacy of Maternal Iron Status Protects against Behavioral, Neuroanatomical, and Growth Deficits in Fetal Alcohol Spectrum Disorders. PLoS ONE 2012, 7, e47499. [Google Scholar] [CrossRef]
- Huebner, S.M.; Tran, T.D.; Rufer, E.S.; Crump, P.M.; Smith, S.M. Maternal Iron Deficiency Worsens the Associative Learning Deficits and Hippocampal and Cerebellar Losses in a Rat Model of Fetal Alcohol Spectrum Disorders. Alcohol. Clin. Exp. Res. 2015, 39, 2097–2107. [Google Scholar] [CrossRef] [PubMed]
- Huebner, S.M.; Blohowiak, S.E.; Kling, P.J.; Smith, S.M. Prenatal Alcohol Exposure Alters Fetal Iron Distribution and Elevates Hepatic Hepcidin in a Rat Model of Fetal Alcohol Spectrum Disorders. J. Nutr. 2016, 146, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.C.; Georgieff, M.K.; Ennis, K.M.; Dodge, N.C.; Wainwright, H.; Meintjes, E.M.; Duggan, C.P.; Molteno, C.D.; Jacobson, J.L.; Jacobson, S.W. Prenatal alcohol-related alterations in maternal, placental, neonatal, and infant iron homeostasis. Am. J. Clin. Nutr. 2021, 114, 1107–1122. [Google Scholar] [CrossRef] [PubMed]
- Sangkhae, V.; Nemeth, E. Regulation of the Iron Homeostatic Hormone Hepcidin. Adv. Nutr. Int. Rev. J. 2017, 8, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Huebner, S.M.; Helfrich, K.; Saini, N.; Blohowiak, S.E.; Cheng, A.A.; Kling, P.J.; Smith, S.M. Dietary Iron Fortification Normalizes Fetal Hematology, Hepcidin, and Iron Distribution in a Rat Model of Prenatal Alcohol Exposure. Alcohol. Clin. Exp. Res. 2018, 42, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The Global Prevalence of Anaemia in 2011; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Mwangi, M.N.; Prentice, A.M.; Verhoef, H. Safety and benefits of antenatal oral iron supplementation in low-income countries: A review. Br. J. Haematol. 2017, 177, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Daru, J.; Cooper, N.A.; Khan, K.S. Systematic review of randomized trials of the effect of iron supplementation on iron stores and oxygen carrying capacity in pregnancy. Acta Obstet. Gynecol. Scand. 2015, 95, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Kashi, B.; Godin, C.M.; Kurzawa, Z.A.; Verney, A.M.J.; Busch-Hallen, J.F.; De-Regil, L.M. Multiple Micronutrient Supplements Are More Cost-effective Than Iron and Folic Acid: Modeling Results from 3 High-Burden Asian Countries. J. Nutr. 2019, 149, 1222–1229. [Google Scholar] [CrossRef]
- Baltussen, R.; Knai, C.; Sharan, M. Iron Fortification and Iron Supplementation are Cost-Effective Interventions to Reduce Iron Deficiency in Four Subregions of the World. J. Nutr. 2004, 134, 2678–2684. [Google Scholar] [CrossRef] [Green Version]
- Georgieff, M.K.; Krebs, N.F.; Cusick, S.E. The Benefits and Risks of Iron Supplementation in Pregnancy and Childhood. Annu. Rev. Nutr. 2019, 39, 121–146. [Google Scholar] [CrossRef]
- Peña-Rosas, J.P.; De-Regil, L.M.; Garcia-Casal, M.N.; Dowswell, T. Daily oral iron supplementation during pregnancy. Cochrane Database Syst. Rev. 2015, 12, CD004736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña-Rosas, J.P.; Viteri, F.E. Effects and safety of preventive oral iron or iron+folic acid supplementation for women during pregnancy. Cochrane Database Syst. Rev. 2009, 4, CD004736. [Google Scholar] [CrossRef]
- Milman, N. Oral Iron Prophylaxis in Pregnancy: Not Too Little and Not Too Much! J. Pregnancy 2012, 2012, 514345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Daily Iron and Folic Acid Supplementation during Pregnancy: Guidance Summary. e-Library of Evidence for Nutrition Actions (eLENA). 2021. Available online: https://www.who.int/elena/titles/guidance_summaries/daily_iron_pregnancy/en/ (accessed on 8 May 2021).
- Saini, N.; Helfrich, K.K.; Kwan, S.T.; Huebner, S.M.; Abazi, J.; Flentke, G.R.; Blohowiak, S.E.; Kling, P.J.; Smith, S.M. Alcohol’s Dysregulation of Maternal–Fetal IL-6 and p-STAT3 Is a Function of Maternal Iron Status. Alcohol. Clin. Exp. Res. 2019, 43, 2332–2343. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.T.C.; Kezer, C.A.; Helfrich, K.K.; Saini, N.; Huebner, S.M.; Flentke, G.R.; Kling, P.J.; Smith, S.M. Maternal iron nutriture modulates placental development in a rat model of fetal alcohol spectrum disorder. Alcohol 2019, 84, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Report of the American Institute of Nutrition Ad Hoc Committee on Standards for Nutritional Studies. J. Nutr. 1977, 107, 1340–1348. [CrossRef]
- Smith, S.M.; Beard, J.L. Norepinephrine turnover in iron deficiency: Effect of two semi-purified diets. Life Sci. 1989, 45, 341–347. [Google Scholar] [CrossRef]
- Minior, V.K.; Levine, B.; Ferber, A.; Guller, S.; Divon, M.Y. Nucleated Red Blood Cells as a Marker of Acute and Chronic Fetal Hypoxia in a Rat Model. Rambam Maimonides Med. J. 2017, 8, e0025. [Google Scholar] [CrossRef] [Green Version]
- Kwan, S.T.; Presswood, B.H.; Helfrich, K.K.; Baulch, J.W.; Mooney, S.M.; Smith, S.M. An interaction between fetal sex and placental weight and efficiency predicts intrauterine growth in response to maternal protein insufficiency and gestational exposure window in a mouse model of FASD. Biol. Sex Differ. 2020, 11, 40. [Google Scholar] [CrossRef]
- Dhakal, P.; Soares, M.J. Single-step PCR-based genetic sex determination of rat tissues and cells. BioTechniques 2017, 62, 232–233. [Google Scholar] [CrossRef] [Green Version]
- Rebouche, C.J.; Wilcox, C.L.; Widness, J. Microanalysis of non-heme iron in animal tissues. J. Biochem. Biophys. Methods 2004, 58, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Hüppi, P.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, K.M.; Haugen, G.; Kiserud, T.; Inskip, H.M.; Cooper, C.; Harvey, N.C.W.; Crozier, S.R.; Robinson, S.M.; Davies, L.; Hanson, M.A. Fetal Liver Blood Flow Distribution: Role in Human Developmental Strategy to Prioritize Fat Deposition versus Brain Development. PLoS ONE 2012, 7, e41759. [Google Scholar] [CrossRef] [Green Version]
- Januszko, O.; Madej, D.; Brzozowska, A.; Kaluza, J. Iron absorption after introducing and discontinuation of iron and zinc supplementation in rats. J. Trace Elements Med. Biol. 2016, 35, 77–82. [Google Scholar] [CrossRef]
- Miller, M.W.; Roskams, A.J.I.; Connor, J.R. Iron Regulation in the Developing Rat Brain: Effect of In Utero Ethanol Exposure. J. Neurochem. 2002, 65, 373–380. [Google Scholar] [CrossRef]
- O’Brien, K.O.; Zavaleta, N.; Caulfield, L.E.; Wen, J.; Abrams, S. Prenatal Iron Supplements Impair Zinc Absorption in Pregnant Peruvian Women. J. Nutr. 2000, 130, 2251–2255. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Ha, J.-H.; Collins, J. Dietary Iron Intake in Excess of Requirements Impairs Intestinal Copper Absorption in Sprague Dawley Rat Dams, Causing Copper Deficiency in Suckling Pups. Biomedicines 2021, 9, 338. [Google Scholar] [CrossRef]
- Katsarou, A.; Pantopoulos, K. Basics and principles of cellular and systemic iron homeostasis. Mol. Asp. Med. 2020, 75, 100866. [Google Scholar] [CrossRef]
- Harrison-Findik, D.D.; Klein, E.; Crist, C.; Evans, J.; Timchenko, N.; Gollan, J. Iron-mediated regulation of liver hepcidin expression in rats and mice is abolished by alcohol. Hepatology 2007, 46, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Costa-Matos, L.; Batista, P.; Monteiro, N.; Simões, M.; Egas, C.; Pereira, J.; Pinho, H.; Santos, N.; Ribeiro, J.; Cipriano, M.A.; et al. Liver hepcidin mRNA expression is inappropriately low in alcoholic patients compared with healthy controls. Eur. J. Gastroenterol. Hepatol. 2012, 24, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, A. Iron, oxidative stress and liver fibrogenesis. J. Hepatol. 1998, 28, 8–13. [Google Scholar] [CrossRef]
- Dey, A.; Cederbaum, A.I. Alcohol and oxidative liver injury. Hepatology 2006, 43, S63–S74. [Google Scholar] [CrossRef]
- Juul, S.E.; Derman, R.J.; Auerbach, M. Perinatal Iron Deficiency: Implications for Mothers and Infants. Neonatology 2019, 115, 269–274. [Google Scholar] [CrossRef]
- Fisher, A.; Nemeth, E. Iron homeostasis during pregnancy. Am. J. Clin. Nutr. 2017, 106 (Suppl. 6), 1567S–1574S. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; The National Academies Press: Washington, DC, USA, 2001; 800p. [Google Scholar]
- Quesenberry, K.E.; Carpenter, J.W. Ferrets, Rabbits, and Rodents: Clinical Medicine and Surgery, 2nd ed.; Saunders: St. Louis, MO, USA, 2004. [Google Scholar]
- Tolkien, Z.; Stecher, L.; Mander, A.P.; Pereira, D.I.A.; Powell, J.J. Ferrous Sulfate Supplementation Causes Significant Gastrointestinal Side-Effects in Adults: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0117383. [Google Scholar] [CrossRef] [Green Version]
- Porta, E.A. Dietary modulation of oxidative stress in alcoholic liver disease in rats. J. Nutr. 1997, 127, 912S–915S. [Google Scholar] [CrossRef] [Green Version]
- Toblli, J.; Cao, G.; Oliveri, L.; Angerosa, M. Effects of Iron Polymaltose Complex, Ferrous Fumarate and Ferrous Sulfate Treatments in Anemic Pregnant Rats, Their Fetuses and Placentas. Inflamm. Allergy-Drug Targets 2013, 12, 190–198. [Google Scholar] [CrossRef]
- Lofruthe, N.; Gallitz, I.; Traeger, L.; Bäumer, N.; Schulze, I.; Kuhlmann, T.; Müller-Tidow, C.; Steinbicker, A.U. Intravenous Iron Carboxymaltose as a Potential Therapeutic in Anemia of Inflammation. PLoS ONE 2016, 11, e0158599. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, X.-M.; Zhang, M.; Yang, C.-Y.; Lv, S.-Y.; Li, S.-F.; Zhong, C.-Y.; Geng, S.-S. Adverse effects of iron deficiency anemia on pregnancy outcome and offspring development and intervention of three iron supplements. Sci. Rep. 2021, 11, 1347. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, N.U.; Cercamondi, C.I.; Brittenham, G.; Zeder, C.; Geurts-Moespot, A.J.; Swinkels, D.W.; Moretti, D.; Zimmermann, M.B. Iron absorption from oral iron supplements given on consecutive versus alternate days and as single morning doses versus twice-daily split dosing in iron-depleted women: Two open-label, randomised controlled trials. Lancet Haematol. 2017, 4, e524–e533. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.; Toblli, J.E.; Romero, J.D.; Monterrosa, B.; Frer, C.; Macagno, E.; Breymann, C. Efficacy and safety of oral iron(III) polymaltose complex versus ferrous sulfate in pregnant women with iron-deficiency anemia: A multicenter, randomized, controlled study. J. Matern. Neonatal Med. 2011, 24, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.M.; Buchanan, G.R.; Adix, L.; Zhang, S.; Gao, A.; McCavit, T.L. Effect of Low-Dose Ferrous Sulfate vs Iron Polysaccharide Complex on Hemoglobin Concentration in Young Children with Nutritional Iron-Deficiency Anemia. JAMA 2017, 317, 2297–2304. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, J. Effects of Ethanol and Maternal Nutritional Status on Fetal Development. Alcohol. Clin. Exp. Res. 1985, 9, 49–55. [Google Scholar] [CrossRef] [PubMed]
- May, P.; Chambers, C.D.; Kalberg, W.O.; Zellner, J.; Feldman, H.; Buckley, D.; Kopald, D.; Hasken, J.M.; Xu, R.; Honerkamp-Smith, G.; et al. Prevalence of Fetal Alcohol Spectrum Disorders in 4 US Communities. JAMA 2018, 319, 474–482. [Google Scholar] [CrossRef]
- May, P.A.; Marais, A.-S.; De Vries, M.M.; Buckley, D.; Kalberg, W.O.; Hasken, J.M.; Stegall, J.M.; Hedrick, D.M.; Robinson, L.K.; Manning, M.A.; et al. The prevalence, child characteristics, and maternal risk factors for the continuum of fetal alcohol spectrum disorders: A sixth population-based study in the same South African community. Drug Alcohol Depend. 2020, 218, 108408. [Google Scholar] [CrossRef]
- May, P.A.; Hamrick, K.J.; Corbin, K.D.; Hasken, J.M.; Marais, A.-S.; Blankenship, J.; Hoyme, H.; Gossage, J.P. Maternal nutritional status as a contributing factor for the risk of fetal alcohol spectrum disorders. Reprod. Toxicol. 2015, 59, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Kable, J.; Coles, C.; Keen, C.; Uriu-Adams, J.; Jones, K.; Yevtushok, L.; Kulikovsky, Y.; Wertelecki, W.; Pedersen, T.; Chambers, C.D. The impact of micronutrient supplementation in alcohol-exposed pregnancies on information processing skills in Ukrainian infants. Alcohol 2015, 49, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Coles, C.D.; Cifasd, T.; Kable, J.A.; Keen, C.L.; Jones, K.L.; Wertelecki, W.; Granovska, I.V.; Pashtepa, A.O.; Chambers, C.D. Dose and Timing of Prenatal Alcohol Exposure and Maternal Nutritional Supplements: Developmental Effects on 6-Month-Old Infants. Matern. Child Health J. 2015, 19, 2605–2614. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | ALC | CON + Iron | ALC + Iron | p Value | |||
|---|---|---|---|---|---|---|---|
| ALC | Iron | ALC × Iron | |||||
| Maternal Outcomes | |||||||
| Age at Breeding (wks) | 10.8 ± 0.6 | 10.2 ± 0.6 | 10.3 ± 0.8 | 10.5 ± 0.9 | Overall p = 0.195 | ||
| Weight on GD0.5 (g) | 236 ± 18 | 226 ± 19 | 230 ± 20 | 232 ± 15 | 0.490 | 0.955 | 0.347 |
| Weight on GD20.5 (g) | 345 ± 18 a | 307 ± 20 c | 339 ± 29 ab | 313 ± 24 bc | <0.001 | 0.962 | 0.442 |
| Gestational weight gain (g) | 109 ± 8 a | 81 ± 25 b | 109 ± 15 ab | 81 ± 28 b | Overall p = 0.003 | ||
| Weight gain GD12.5–20.5 (g) | 50 ± 6 a | 24 ± 24 b | 54 ± 7 a | 25 ± 22 b | Overall p < 0.001 | ||
| Non-pregnancy weight (g) | 294 ± 20 a | 258 ± 16 b | 284 ± 24 ab | 269 ± 18 ab | <0.001 | 0.967 | 0.117 |
| Gestational food consumption (g) | 352 ± 22 a | 306 ± 31 b | 341 ± 35 ab | 320 ± 41 ab | 0.003 | 0.890 | 0.248 |
| Food conversion ratio (kcal/g) | 12.9 ± 1.0 a | 16.42 ± 4.1 b | 12.6 ± 0.8 a | 18.6 ± 10.1 b | Overall p = 0.001 | ||
| Litter Outcomes | |||||||
| Litter Size (n) | 10.0 ± 1.8 | 11.2 ± 1.1 | 10.9 ± 1.7 | 9.8 ± 2.8 | Overall p = 0.477 | ||
| Percent Survival (%) | 96 ± 9 | 93 ± 8 | 96 ± 5 | 87 ± 12 | Overall p = 0.332 | ||
| Percent Male (%) | 54 ± 20 | 47 ± 14 | 60 ± 7 | 45 ± 20 | Overall p = 0.323 | ||
| Uterine Horn Weight (g) | 51.0 ± 7.1 ab | 48.6 ± 6.9 ab | 55.0 ± 8.6 a | 43.3 ± 11.8 b | 0.024 | 0.830 | 0.127 |
| Uterine Weight Per Fetus (g) | 5.1 ± 0.4 a | 4.3 ± 0.4 b | 5.1 ± 0.3 a | 4.5 ± 0.5 b | <0.001 | 0.767 | 0.426 |
| MALES | p Value | ||||||
|---|---|---|---|---|---|---|---|
| CON | ALC | CON + Iron | ALC + Iron | ALC | Iron | ALC × Iron | |
| Absolute weight (g) | |||||||
| Fetus | 3.69 ± 0.28 a | 3.03 ± 0.41 b | 3.70 ± 0.28 a | 3.22 ± 0.42 b | <0.001 | 0.449 | 0.341 |
| Placenta | 0.39 ± 0.07 a | 0.35 ± 0.05 bc | 0.38 ± 0.06 ab | 0.34 ± 0.04 c | 0.012 | 0.391 | 0.497 |
| Brain | 0.16 ± 0.02 a | 0.14 ± 0.01 b | 0.15 ± 0.02 a | 0.15 ± 0.02 ab | 0.004 | 0.725 | 0.014 |
| Liver | 0.26 ± 0.05 a | 0.21 ± 0.04 b | 0.26 ± 0.04 a | 0.22 ± 0.05 b | <0.001 | 0.960 | 0.392 |
| Heart | 0.018 ± 0.002 a | 0.015 ± 0.002 b | 0.018 ± 0.002 a | 0.016 ± 0.003 ab | 0.006 | 0.668 | 0.416 |
| Placental efficiency | 9.66 ± 1.43 a | 8.70 ± 1.66 b | 9.89 ± 1.21 a | 9.28 ± 1.22 ab | 0.043 | 0.232 | 0.921 |
| Relative weight (g/100 g total body weight) | |||||||
| Brain | 4.3 ± 0.5 a | 4.8 ± 0.7 b | 4.1 ± 0.4 a | 4.7 ± 0.6 b | <0.001 | 0.408 | 0.593 |
| Liver | 7.1 ± 1.3 | 6.8 ± 1.0 | 7.0 ± 1.0 | 6.9 ± 0.9 | 0.270 | 0.824 | 0.840 |
| Heart | 0.47 ± 0.06 a | 0.50 ± 0.04 bc | 0.48 ± 0.05 ab | 0.51 ± 0.05 c | 0.002 | 0.802 | 0.884 |
| Brain to liver ratio | 0.61 ± 0.11 a | 0.72 ± 0.17 b | 0.60 ± 0.11 a | 0.70 ± 0.14 b | <0.001 | 0.833 | 0.969 |
| FEMALES | p Value | ||||||
|---|---|---|---|---|---|---|---|
| CON | ALC | CON + Iron | ALC + Iron | ALC | Iron | ALC × Iron | |
| Absolute weight (g) | |||||||
| Fetus | 3.52 ± 0.30 a | 2.88 ± 0.41 b | 3.46 ± 0.27 a | 2.94 ± 0.48 b | <0.001 | 0.819 | 0.562 |
| Placenta | 0.37 ± 0.07 | 0.34 ± 0.04 | 0.35 ± 0.05 | 0.32 ± 0.04 | 0.163 | 0.322 | 0.809 |
| Brain | 0.16 ± 0.02 a | 0.14 ± 0.01 b | 0.15 ± 0.02 ab | 0.15 ± 0.01 b | 0.044 | 0.518 | 0.062 |
| Liver | 0.27 ± 0.05 a | 0.20 ± 0.04 b | 0.25 ± 0.03 a | 0.21 ± 0.04 b | <0.001 | 0.588 | 0.293 |
| Heart | 0.016 ± 0.003 a | 0.015 ± 0.002 c | 0.016 ± 0.002 ab | 0.015 ± 0.003 bc | 0.019 | 0.624 | 0.607 |
| Placental efficiency | 9.73 ± 1.51 ab | 8.69 ± 1.25 c | 10.11 ± 1.21 a | 9.04 ± 1.35 bc | 0.006 | 0.327 | 0.894 |
| Relative weight (g/100 g total body weight) | |||||||
| Brain | 4.5 ± 0.4 a | 5.1 ± 0.7 b | 4.3 ± 0.5 a | 5.1 ± 0.7 b | <0.001 | 0.619 | 0.654 |
| Liver | 7.6 ± 1.0 a | 7.1 ± 1.0 b | 7.2 ± 0.6 ab | 7.0 ± 0.8 b | 0.039 | 0.147 | 0.351 |
| Heart | 0.47 ± 0.06 a | 0.51 ± 0.05 b | 0.47 ± 0.05 a | 0.51 ± 0.07 b | 0.003 | 0.904 | 0.745 |
| Brain to liver ratio | 0.59 ± 0.10 a | 0.74 ± 0.16 b | 0.60 ± 0.09 a | 0.75 ± 0.16 b | <0.001 | 0.905 | 0.937 |
| CON | ALC | CON + Iron | ALC + Iron | p Value | |||
|---|---|---|---|---|---|---|---|
| ALC | Iron | ALC × Iron | |||||
| RBC (×106 cells/μL) | 6.55 ± 0.26 | 6.52 ± 0.23 | 6.60 ± 0.19 | 6.61 ± 0.45 | 0.878 | 0.502 | 0.856 |
| Hemoglobin (g/dL) | 12.4 ± 0.5 | 11.9 ± 0.4 | 12.0 ± 0.3 | 12.2 ± 0.7 | 0.478 | 0.866 | 0.071 |
| Hematocrit (%) | 36.8 ± 1.5 | 35.5 ± 0.8 | 36.4 ± 0.7 | 36.6 ± 2.1 | 0.254 | 0.508 | 0.174 |
| MCV (fL/cell) | 56.1 ± 1.1 | 54.6 ± 1.2 | 55.2 ± 1.1 | 55.3 ± 1.5 | 0.108 | 0.813 | 0.051 |
| MCH (pg/cell) | 18.9 ± 0.5 a | 18.3 ± 0.4 ab | 18.2 ± 0.5 b | 18.5 ± 0.6 ab | 0.368 | 0.180 | 0.014 |
| MCHC (pg/dL) | 33.6 ± 0.4 a | 33.5 ± 0.4 a | 33.0 ± 0.6 a | 33.4 ± 0.5 a | 0.386 | 0.038 | 0.214 |
| PLT (×103 cells/μL) | 859 ± 149 | 788 ± 118 | 881 ± 138 | 848 ± 130 | 0.265 | 0.375 | 0.677 |
| RDW-CV (%) | 13.0 ± 0.3 | 13.0 ± 0.4 | 12.9 ± 0.4 | 13.1 ± 0.5 | 0.493 | 0.930 | 0.594 |
| MPV (fL/cell) | 7.8 ± 0.4 | 7.7 ± 0.3 | 7.7 ± 0.2 | 7.6 ± 0.2 | Overall p = 0.452 | ||
| ZnPP (μmol ZnPP/mol heme) | 69.7 ± 5.2 | 65.2 ± 4.0 | 65.3 ± 3.6 | 66.7 ± 5.0 | 0.325 | 0.345 | 0.063 |
| nRBC (per 100 WBCs) | 204 ± 38 a | 157 ± 44 ab | 150 ± 57 ab | 100 ± 45 b | 0.004 | 0.001 | 0.902 |
| CON | ALC | CON + Iron | ALC + Iron | p Value | |||
|---|---|---|---|---|---|---|---|
| ALC | Iron | ALC × Iron | |||||
| RBC (×106 cells/μL) | 1.65 ± 0.13 ab | 1.47 ± 0.18 b | 1.67 ± 0.12 a | 1.64 ± 0.14 ab | 0.039 | 0.058 | 0.141 |
| Hemoglobin (g/dL) | 10.2 ± 0.6 a | 10.0 ± 0.5 a | 10.5 ± 0.4 a | 10.5 ± 0.6 a | 0.596 | 0.044 | 0.490 |
| Hematocrit (%) | 31.4 ± 2.1 ab | 28.5 ± 3.0 b | 31.8 ± 1.7 a | 31.2 ± 2.3 ab | 0.037 | 0.056 | 0.162 |
| MCV (fL/cell) | 190.6 ± 3.3 | 194.4 ± 4.5 | 190.7 ± 3.5 | 190.6 ± 2.8 | 0.138 | 0.138 | 0.123 |
| MCH (pg/cell) | 62.2 ± 4.6 | 69.0 ± 10.5 | 62.8 ± 5.3 | 64.1 ± 4.3 | Overall p = 0.225 | ||
| MCHC (pg/dL) | 32.6 ± 1.9 | 35.4 ± 4.5 | 32.9 ± 2.2 | 33.6 ± 1.9 | Overall p = 0.273 | ||
| PLT (×103 cells/μL) | 135.4 ± 20.3 | 122.7 ± 15.7 | 140.4 ± 23.9 | 130.9 ± 23.2 | 0.126 | 0.360 | 0.818 |
| RDW-CV (%) | 14.9 ± 0.8 ab | 14.1 ± 1.1 b | 15.1 ± 0.7 ab | 15.6 ± 0.9 a | 0.541 | 0.012 | 0.052 |
| MPV (fL/cell) | 12.0 ± 0.5 a | 12.4 ± 0.4 a | 11.9 ± 0.3 a | 12.2 ± 0.3 a | 0.011 | 0.061 | 0.565 |
| ZnPP (μmol ZnPP/mol heme) | 65.4 ± 8.7 a | 59.0 ± 4.2 a | 59.2 ± 3.7 a | 61.9 ± 6.4 a | 0.396 | 0.479 | 0.043 |
| nRBC (per 100 WBCs) | 907 ± 397 a | 1153 ± 241 ab | 768 ± 393 a | 1985 ± 1384 b | 0.001 | 0.556 | 0.102 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helfrich, K.K.; Saini, N.; Kwan, S.T.C.; Rivera, O.C.; Hodges, R.; Smith, S.M. Gestational Iron Supplementation Improves Fetal Outcomes in a Rat Model of Prenatal Alcohol Exposure. Nutrients 2022, 14, 1653. https://doi.org/10.3390/nu14081653
Helfrich KK, Saini N, Kwan STC, Rivera OC, Hodges R, Smith SM. Gestational Iron Supplementation Improves Fetal Outcomes in a Rat Model of Prenatal Alcohol Exposure. Nutrients. 2022; 14(8):1653. https://doi.org/10.3390/nu14081653
Chicago/Turabian StyleHelfrich, Kaylee K., Nipun Saini, Sze Ting Cecilia Kwan, Olivia C. Rivera, Rachel Hodges, and Susan M. Smith. 2022. "Gestational Iron Supplementation Improves Fetal Outcomes in a Rat Model of Prenatal Alcohol Exposure" Nutrients 14, no. 8: 1653. https://doi.org/10.3390/nu14081653