
Sex-Related Differences in Voluntary Alcohol Intake and mRNA Coding for Synucleins in the Brain of Adult Rats Prenatally Exposed to Alcohol

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
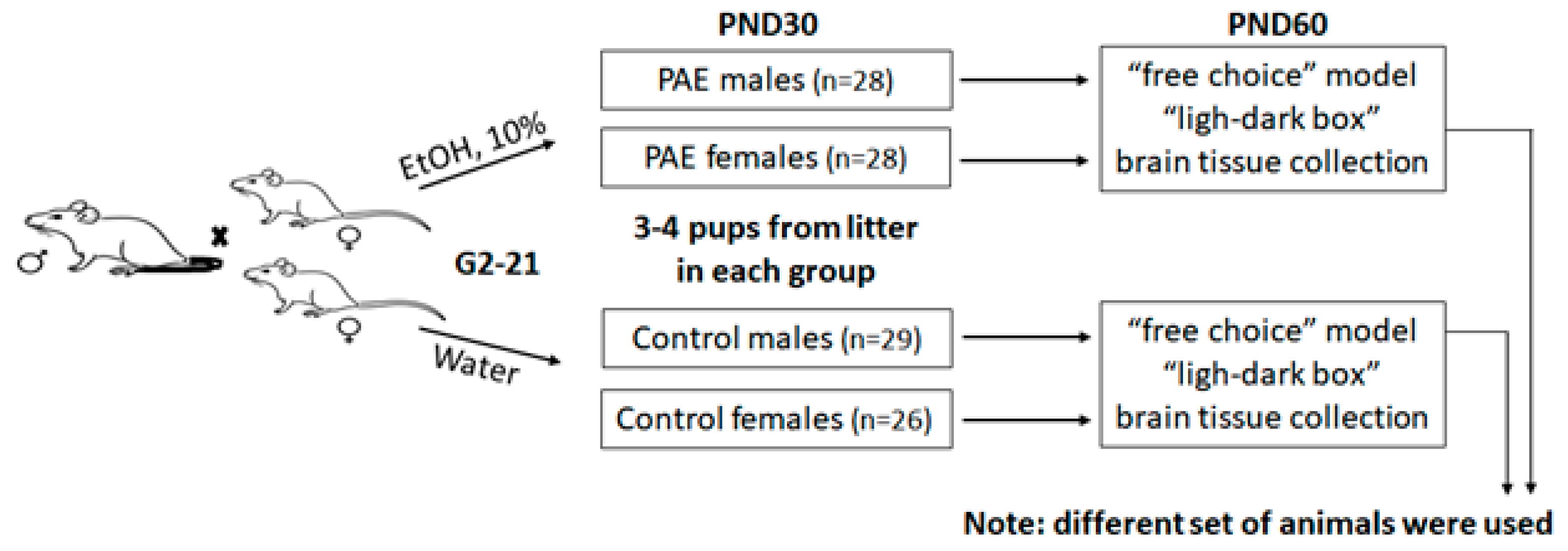
2.1. Animals
2.2. Two-Bottle “10% Alcohol vs. Water” Choice Drinking Paradigm (Voluntary Alcohol Consumption)
2.3. Light-Dark Box
2.4. Tissue Collection
2.5. RNA Extraction, cDNA Synthesis and Quantitative RT-PCR
2.6. Data Analysis
3. Results
3.1. Observation of Pregnant Dam and Offspring
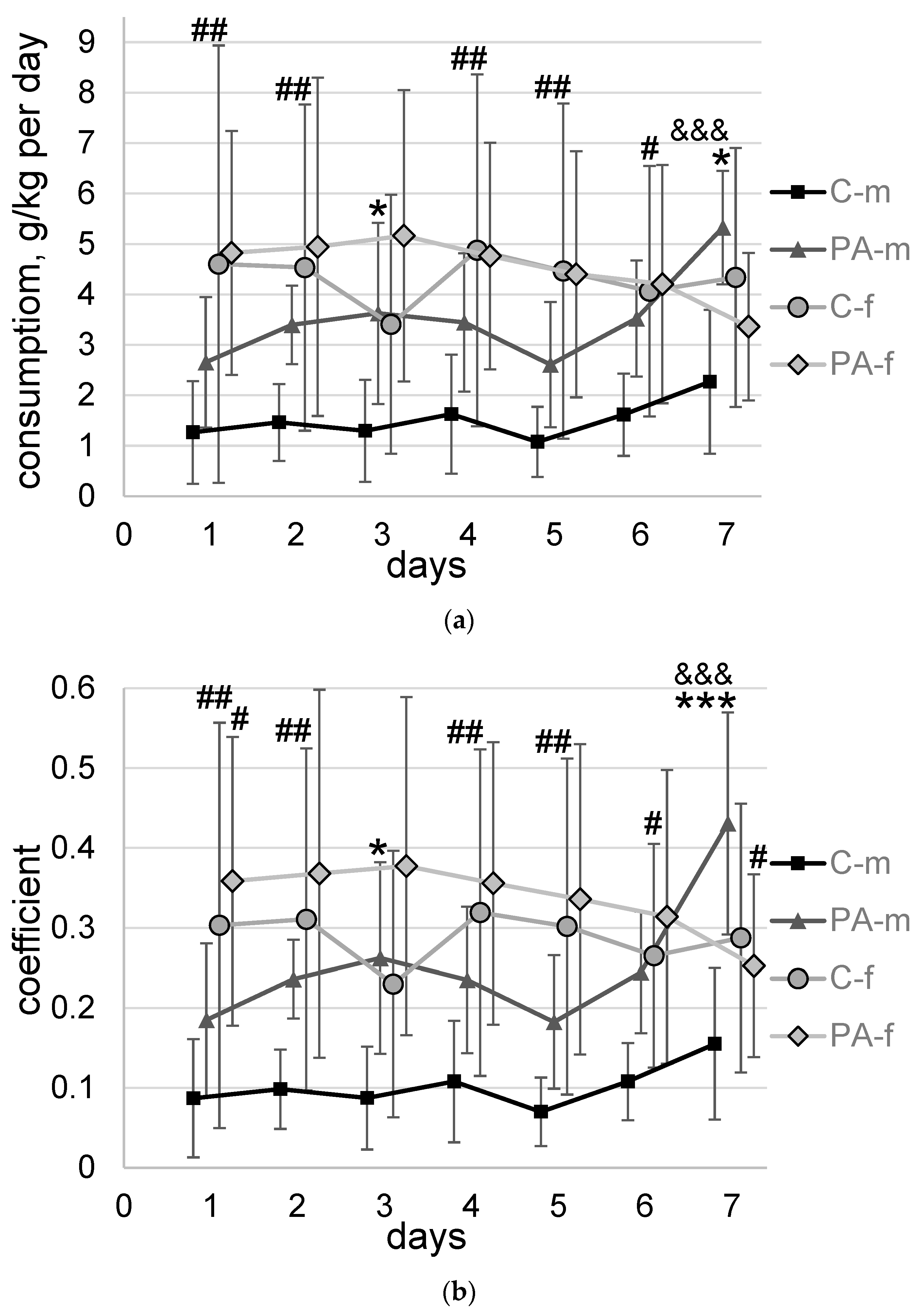
3.2. Voluntary Alcohol Consumption
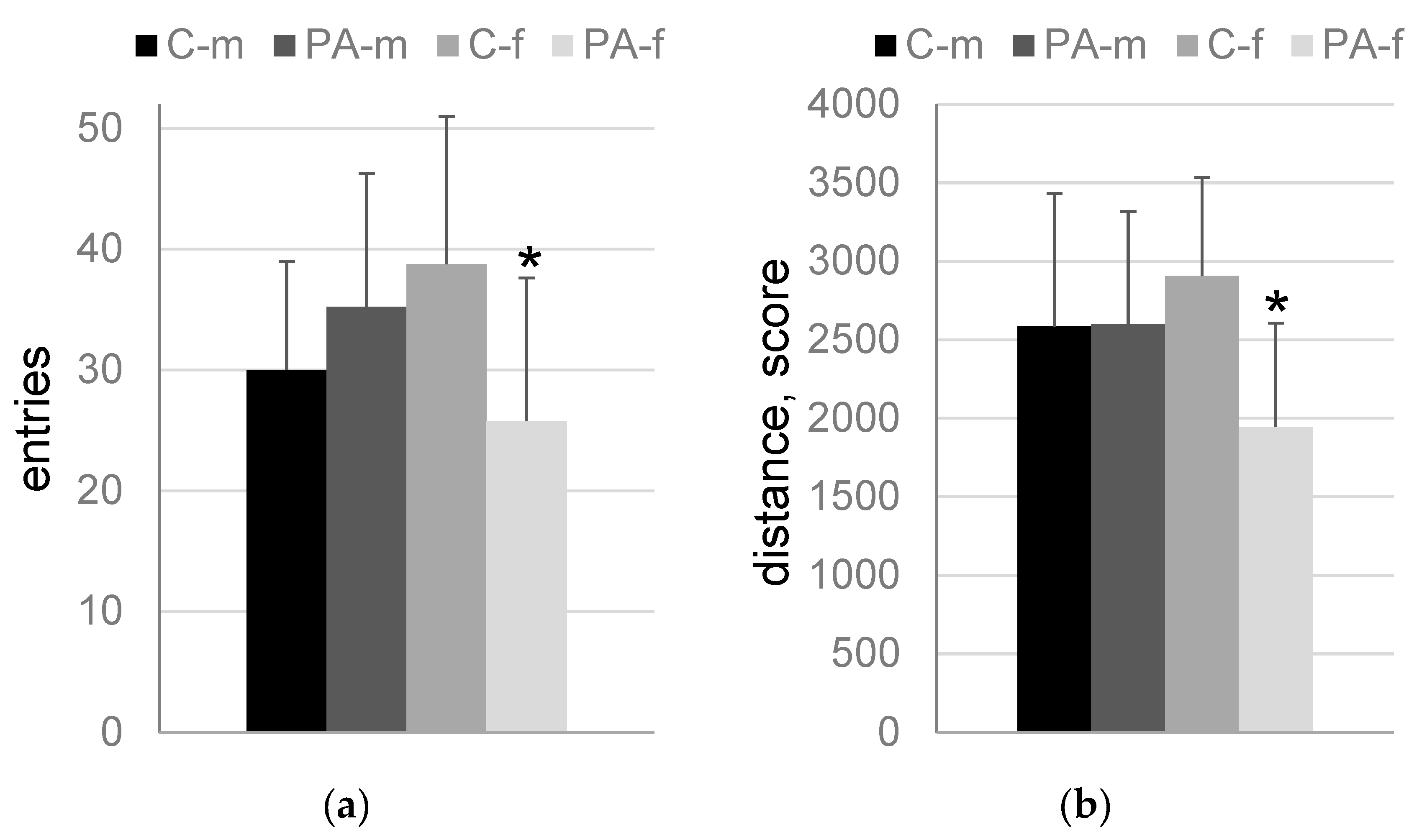
3.3. The “Light-Dark Box” Test
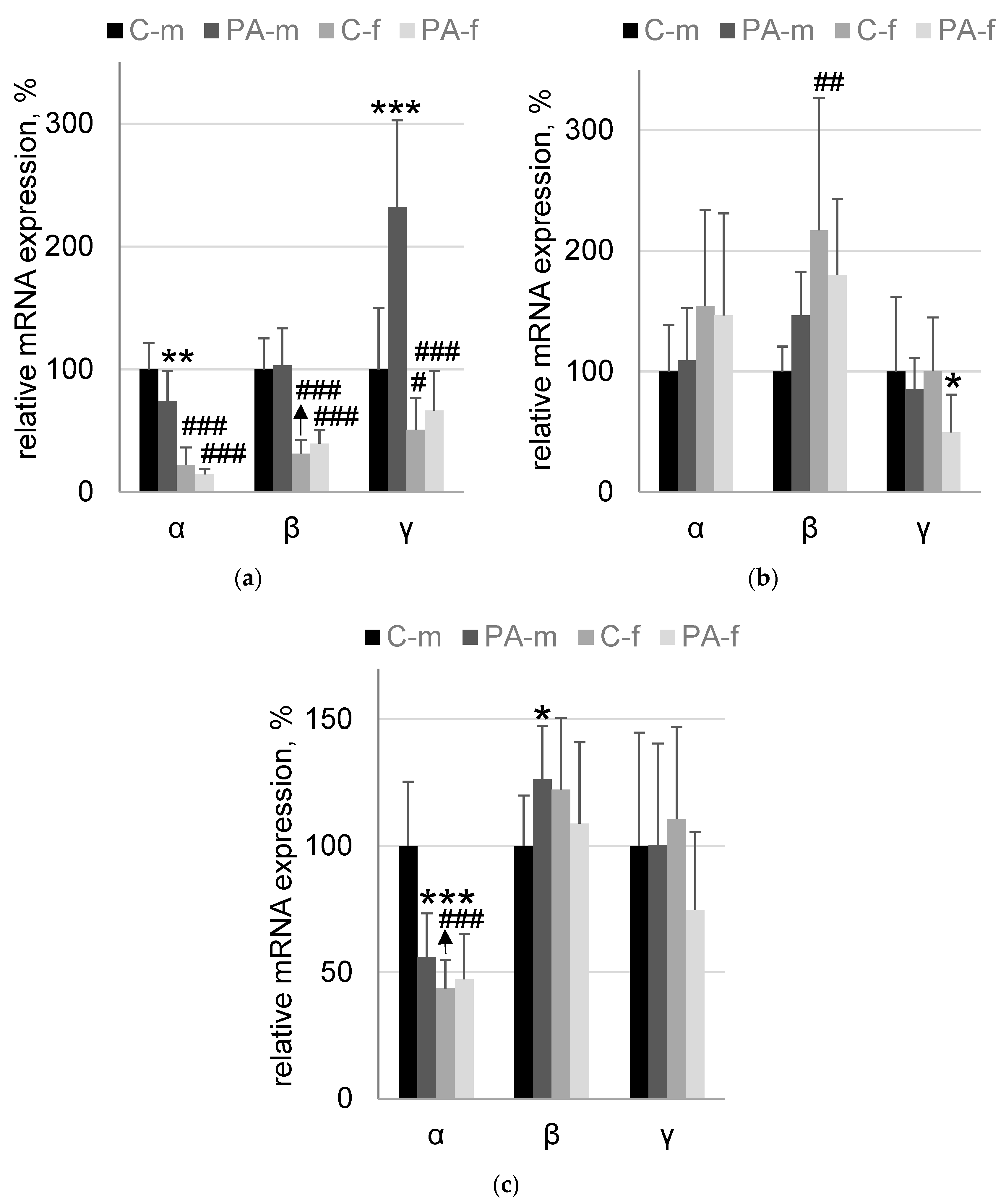
3.4. Synucleins mRNA Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, I.; Dandavate, R.; Gupta, P.; Agrawal, V.; Kapoor, M. Recent advances in genetic studies of alcohol use disorders. Curr. Genet. Med. Rep. 2020, 8, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Longley, M.J.; Lee, J.; Jung, J.; Lohoff, F.W. Epigenetics of alcohol use disorder-A review of recent advances in DNA methylation profiling. Addict. Biol. 2021, 26, e13006. [Google Scholar] [CrossRef] [PubMed]
- Gaztanaga, M.; Angulo-Alcalde, A.; Chotro, M.G. Prenatal Alcohol Exposure as a Case of Involuntary Early Onset of Alcohol Use: Consequences and Proposed Mechanisms From Animal Studies. Front. Behav. Neurosci. 2020, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.L.; Akkaya-Hocagil, T.; Ryan, L.M.; Dodge, N.C.; Richardson, G.A.; Olson, H.C.; Coles, C.D.; Day, N.L.; Cook, R.J.; Jacobson, S.W. Effects of prenatal alcohol exposure on cognitive and behavioral development: Findings from a hierarchical meta-analysis of data from six prospective longitudinal U.S. cohorts. Alcohol. Clin. Exp. Res. 2021, 45, 2040–2058. [Google Scholar] [CrossRef]
- Charness, M.E. Fetal Alcohol Spectrum Disorders: Awareness to Insight in Just 50 Years. Alcohol Res. 2022, 42, 05. [Google Scholar] [CrossRef]
- Das, J. SNARE Complex-Associated Proteins and Alcohol. Alcohol. Clin. Exp. Res. 2020, 44, 7–18. [Google Scholar] [CrossRef]
- Nemani, V.M.; Lu, W.; Berge, V.; Nakamura, K.; Onoa, B.; Lee, M.K.; Chaudhry, F.A.; Nicoll, R.A.; Edwards, R.H. Increased expression of alpha-synuclein reduces neurotransmitter release by inhibiting synaptic vesicle reclustering after endocytosis. Neuron 2010, 65, 66–79. [Google Scholar] [CrossRef]
- Cahill, C.M.; Aleyadeh, R.; Gao, J.; Wang, C.; Rogers, J.T. Alpha-Synuclein in Alcohol Use Disorder, Connections with Parkinson’s Disease and Potential Therapeutic Role of 5’ Untranslated Region-Directed Small Molecules. Biomolecules 2020, 10, 1465. [Google Scholar] [CrossRef]
- Levey, D.F.; Le-Niculescu, H.; Frank, J.; Ayalew, M.; Jain, N.; Kirlin, B.; Learman, R.; Winiger, E.; Rodd, Z.; Shekhar, A.; et al. Genetic risk prediction and neurobiological understanding of alcoholism. Transl. Psychiatry 2014, 4, e391. [Google Scholar] [CrossRef]
- Janeczek, P.; MacKay, R.K.; Lea, R.A.; Dodd, P.R.; Lewohl, J.M. Reduced expression of alpha-synuclein in alcoholic brain: Influence of SNCA-Rep1 genotype. Addict. Biol. 2014, 19, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Bonsch, D.; Reulbach, U.; Bayerlein, K.; Hillemacher, T.; Kornhuber, J.; Bleich, S. Elevated alpha synuclein mRNA levels are associated with craving in patients with alcoholism. Biol. Psychiatry 2004, 56, 984–986. [Google Scholar] [CrossRef] [PubMed]
- Foroud, T.; Wetherill, L.F.; Liang, T.; Dick, D.M.; Hesselbrock, V.; Kramer, J.; Nurnberger, J.; Schuckit, M.; Carr, L.; Porjesz, B.; et al. Association of alcohol craving with alpha-synuclein (SNCA). Alcohol. Clin. Exp. Res. 2007, 31, 537–545. [Google Scholar] [CrossRef]
- Rotermund, C.; Reolon, G.K.; Leixner, S.; Boden, C.; Bilbao, A.; Kahle, P.J. Enhanced motivation to alcohol in transgenic mice expressing human alpha-synuclein. J. Neurochem. 2017, 143, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Kimpel, M.W.; McClintick, J.N.; Skillman, A.R.; McCall, K.; Edenberg, H.J.; Carr, L.G. Candidate genes for alcohol preference identified by expression profiling in alcohol-preferring and -nonpreferring reciprocal congenic rats. Genome Biol. 2010, 11, R11. [Google Scholar] [CrossRef] [PubMed]
- Janowska, M.K.; Wu, K.P.; Baum, J. Unveiling transient protein-protein interactions that modulate inhibition of alpha-synuclein aggregation by beta-synuclein, a pre-synaptic protein that co-localizes with alpha-synuclein. Sci. Rep. 2015, 5, 15164. [Google Scholar] [CrossRef]
- Buchman, V.L.; Hunter, H.J.; Pinon, L.G.; Thompson, J.; Privalova, E.M.; Ninkina, N.N.; Davies, A.M. Persyn, a member of the synuclein family, has a distinct pattern of expression in the developing nervous system. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 9335–9341. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Kokhan, T.Y.G.; Samsonova, A.N.; Fisenko, V.P.; Ustyugov, A.A.; Aliev, G. The Dopaminergic Dysfunction and Altered Working Memory Performance of Aging Mice Lacking Gamma-synuclein Gene. CNS Neurol. Disord. Drug Targets 2018, 17, 604–607. [Google Scholar] [CrossRef]
- Senior, S.L.; Ninkina, N.; Deacon, R.; Bannerman, D.; Buchman, V.L.; Cragg, S.J.; Wade-Martins, R. Increased striatal dopamine release and hyperdopaminergic-like behaviour in mice lacking both alpha-synuclein and gamma-synuclein. Eur. J. Neurosci. 2008, 27, 947–957. [Google Scholar] [CrossRef]
- Wise, R.A.; Jordan, C.J. Dopamine, behavior, and addiction. J. Biomed. Sci. 2021, 28, 83. [Google Scholar] [CrossRef]
- Nutt, D.; Hayes, A.; Fonville, L.; Zafar, R.; Palmer, E.O.C.; Paterson, L.; Lingford-Hughes, A. Alcohol and the Brain. Nutrients 2021, 13, 3938. [Google Scholar] [CrossRef]
- White, A.M. Gender Differences in the Epidemiology of Alcohol Use and Related Harms in the United States. Alcohol Res. 2020, 40, 01. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, M.G.; Patalay, P.; Mackey, S.; Allen, N.B.; Batalla, A.; Bellani, M.; Chye, Y.; Cousijn, J.; Goudriaan, A.E.; Hester, R.; et al. Gender-related neuroanatomical differences in alcohol dependence: Findings from the ENIGMA Addiction Working Group. Neuroimage Clin. 2021, 30, 102636. [Google Scholar] [CrossRef] [PubMed]
- Flores-Bonilla, A.; De Oliveira, B.; Silva-Gotay, A.; Lucier, K.W.; Richardson, H.N. Shortening time for access to alcohol drives up front-loading behavior, bringing consumption in male rats to the level of females. Biol. Sex Differ. 2021, 12, 51. [Google Scholar] [CrossRef]
- Datta, U.; Schoenrock, S.E.; Bubier, J.A.; Bogue, M.A.; Jentsch, J.D.; Logan, R.W.; Tarantino, L.M.; Chesler, E.J. Prospects for finding the mechanisms of sex differences in addiction with human and model organism genetic analysis. Genes Brain Behav. 2020, 19, e12645. [Google Scholar] [CrossRef]
- Patten, A.R.; Fontaine, C.J.; Christie, B.R. A comparison of the different animal models of fetal alcohol spectrum disorders and their use in studying complex behaviors. Front. Pediatr. 2014, 2, 93. [Google Scholar] [CrossRef]
- West, J.R. Fetal alcohol-induced brain damage and the problem of determining temporal vulnerability: A review. Alcohol Drug Res. 1987, 7, 423–441. [Google Scholar]
- Quinn, R. Comparing rat’s to human’s age: How old is my rat in people years? Nutrition 2005, 21, 775–777. [Google Scholar] [CrossRef]
- Baer, J.S.; Sampson, P.D.; Barr, H.M.; Connor, P.D.; Streissguth, A.P. A 21-year longitudinal analysis of the effects of prenatal alcohol exposure on young adult drinking. Arch. Gen. Psychiatry 2003, 60, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.; Andreu-Fernandez, V.; Navarro-Tapia, E.; Aras-Lopez, R.; Serra-Delgado, M.; Martinez, L.; Garcia-Algar, O.; Gomez-Roig, M.D. Murine Models for the Study of Fetal Alcohol Spectrum Disorders: An Overview. Front. Pediatr. 2020, 8, 359. [Google Scholar] [CrossRef]
- Youngentob, S.L.; Glendinning, J.I. Fetal ethanol exposure increases ethanol intake by making it smell and taste better. Proc. Natl. Acad. Sci. USA 2009, 106, 5359–5364. [Google Scholar] [CrossRef]
- Glendinning, J.I.; Simons, Y.M.; Youngentob, L.; Youngentob, S.L. Fetal ethanol exposure attenuates aversive oral effects of TrpV1, but not TrpA1 agonists in rats. Exp. Biol. Med. 2012, 237, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, P.; Han, X.; Zuo, W.; Mei, Q.; Bian, E.Y.; Umeugo, J.; Ye, J. Differences between male and female rats in alcohol drinking, negative affects and neuronal activity after acute and prolonged abstinence. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 163–176. [Google Scholar]
- Blanchard, B.A.; Glick, S.D. Sex differences in mesolimbic dopamine responses to ethanol and relationship to ethanol intake in rats. Recent Dev. Alcohol. 1995, 12, 231–241. [Google Scholar] [CrossRef]
- Spence, J.P.; Reiter, J.L.; Qiu, B.; Gu, H.; Garcia, D.K.; Zhang, L.; Graves, T.; Williams, K.E.; Bice, P.J.; Zou, Y.; et al. Estrogen-Dependent Upregulation of Adcyap1r1 Expression in Nucleus Accumbens Is Associated With Genetic Predisposition of Sex-Specific QTL for Alcohol Consumption on Rat Chromosome 4. Front. Genet. 2018, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, M.A.S.; Pinho, R.; Paiva, I.; Outeiro, T.F. The yin and yang of alpha-synuclein-associated epigenetics in Parkinson’s disease. Brain A J. Neurol. 2017, 140, 878–886. [Google Scholar] [CrossRef]
- Desplats, P.; Spencer, B.; Coffee, E.; Patel, P.; Michael, S.; Patrick, C.; Adame, A.; Rockenstein, E.; Masliah, E. Alpha-synuclein sequesters Dnmt1 from the nucleus: A novel mechanism for epigenetic alterations in Lewy body diseases. J. Biol. Chem. 2011, 286, 9031–9037. [Google Scholar] [CrossRef] [PubMed]
- Razumkina, E.; Anokhin, P.; Sarycheva, N.; Shamakina, I. Prenatal alcohol exposure increases DNA-methyltransferases 1 and 3a its mRNA levels in the rat mesolimbic brain areas. Eur. Neuropsychopharmacol. 2019, 29, S312–S313. [Google Scholar] [CrossRef]
- Gorbatyuk, O.S.; Li, S.; Nash, K.; Gorbatyuk, M.; Lewin, A.S.; Sullivan, L.F.; Mandel, R.J.; Chen, W.; Meyers, C.; Manfredsson, F.P.; et al. In vivo RNAi-mediated alpha-synuclein silencing induces nigrostriatal degeneration. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 1450–1457. [Google Scholar] [CrossRef]
- McCormack, A.L.; Mak, S.K.; Henderson, J.M.; Bumcrot, D.; Farrer, M.J.; Di Monte, D.A. Alpha-synuclein suppression by targeted small interfering RNA in the primate substantia nigra. PLoS ONE 2010, 5, e12122. [Google Scholar] [CrossRef]
- Polissidis, A.; Koronaiou, M.; Kollia, V.; Koronaiou, E.; Nakos-Bimpos, M.; Bogiongko, M.; Vrettou, S.; Karali, K.; Casadei, N.; Riess, O.; et al. Psychosis-Like Behavior and Hyperdopaminergic Dysregulation in Human alpha-Synuclein BAC Transgenic Rats. Mov. Disord. 2021, 36, 716–728. [Google Scholar] [CrossRef]
- Hirth, N.; Meinhardt, M.W.; Noori, H.R.; Salgado, H.; Torres-Ramirez, O.; Uhrig, S.; Broccoli, L.; Vengeliene, V.; Rossmanith, M.; Perreau-Lenz, S.; et al. Convergent evidence from alcohol-dependent humans and rats for a hyperdopaminergic state in protracted abstinence. Proc. Natl. Acad. Sci. USA 2016, 113, 3024–3029. [Google Scholar] [CrossRef] [PubMed]
- Hansson, A.C.; Grunder, G.; Hirth, N.; Noori, H.R.; Spanagel, R.; Sommer, W.H. Dopamine and opioid systems adaptation in alcoholism revisited: Convergent evidence from positron emission tomography and postmortem studies. Neurosci. Biobehav. Rev. 2019, 106, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.R.; Mulholland, P.J.; Truitt, W.A.; Waeiss, R.A.; Engleman, E.A.; Bell, R.L.; Rodd, Z.A. Adolescent Intermittent Ethanol (AIE) Enhances the Dopaminergic Response to Ethanol within the Mesolimbic Pathway during Adulthood: Alterations in Cholinergic/Dopaminergic Genes Expression in the Nucleus Accumbens Shell. Int. J. Mol. Sci. 2021, 22, 11733. [Google Scholar] [CrossRef]
- Santangelo, V.; Cavallina, C.; Colucci, P.; Santori, A.; Macri, S.; McGaugh, J.L.; Campolongo, P. Enhanced brain activity associated with memory access in highly superior autobiographical memory. Proc. Natl. Acad. Sci. USA 2018, 115, 7795–7800. [Google Scholar] [CrossRef] [PubMed]
- Wilhoit, L.F.; Scott, D.A.; Simecka, B.A. Fetal Alcohol Spectrum Disorders: Characteristics, Complications, and Treatment. Community Ment Health J. 2017, 53, 711–718. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Zhang, T. Spatial cognition and sexually dimorphic synaptic plasticity balance impairment in rats with chronic prenatal ethanol exposure. Behav. Brain Res. 2013, 256, 564–574. [Google Scholar] [CrossRef]
- An, L.; Zhang, T. Prenatal ethanol exposure impairs spatial cognition and synaptic plasticity in female rats. Alcohol 2015, 49, 581–588. [Google Scholar] [CrossRef]
- Guerri, C.; Bazinet, A.; Riley, E.P. Foetal Alcohol Spectrum Disorders and alterations in brain and behaviour. Alcohol Alcohol. 2009, 44, 108–114. [Google Scholar] [CrossRef]
- Bon, L.I.; Zimatkin, S.M. Disruption of synaptogenesis in the rats brain cortex after antenatal alcoholisation. J. Grodno State Med. Univ. 2017, 15, 538–543. [Google Scholar] [CrossRef]
- Sadrian, B.; Lopez-Guzman, M.; Wilson, D.A.; Saito, M. Distinct neurobehavioral dysfunction based on the timing of developmental binge-like alcohol exposure. Neuroscience 2014, 280, 204–219. [Google Scholar] [CrossRef]
- Hsu, L.J.; Mallory, M.; Xia, Y.; Veinbergs, I.; Hashimoto, M.; Yoshimoto, M.; Thal, L.J.; Saitoh, T.; Masliah, E. Expression pattern of synucleins (non-Abeta component of Alzheimer’s disease amyloid precursor protein/alpha-synuclein) during murine brain development. J. Neurochem. 1998, 71, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.D.; Rueter, S.M.; Trojanowski, J.Q.; Lee, V.M. Synucleins are developmentally expressed, and alpha-synuclein regulates the size of the presynaptic vesicular pool in primary hippocampal neurons. J. Neurosci. 2000, 20, 3214–3220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Watanabe, Y.; Tsujimura, A.; Tatebe, H.; Miyata, S.; Tokuda, T.; Mizuno, T.; Tanaka, M. Differential expression of alpha-synuclein in hippocampal neurons. PLoS ONE 2014, 9, e89327. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Watanabe, Y.; Tsujimura, A.; Tanaka, M. Expression of alpha-synuclein is regulated in a neuronal cell type-dependent manner. Anat. Sci. Int. 2019, 94, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Gureviciene, I.; Gurevicius, K.; Tanila, H. Role of alpha-synuclein in synaptic glutamate release. Neurobiol. Dis. 2007, 28, 83–89. [Google Scholar] [CrossRef]
- Cheng, H.; Kellar, D.; Lake, A.; Finn, P.; Rebec, G.V.; Dharmadhikari, S.; Dydak, U.; Newman, S. Effects of Alcohol Cues on MRS Glutamate Levels in the Anterior Cingulate. Alcohol Alcohol. 2018, 53, 209–215. [Google Scholar] [CrossRef]
- Ceccarini, J.; Leurquin-Sterk, G.; Crunelle, C.L.; de Laat, B.; Bormans, G.; Peuskens, H.; Van Laere, K. Recovery of Decreased Metabotropic Glutamate Receptor 5 Availability in Abstinent Alcohol-Dependent Patients. J. Nucl. Med. 2020, 61, 256–262. [Google Scholar] [CrossRef]
- Burnette, E.M.; Nieto, S.J.; Grodin, E.N.; Meredith, L.R.; Hurley, B.; Miotto, K.; Gillis, A.J.; Ray, L.A. Novel Agents for the Pharmacological Treatment of Alcohol Use Disorder. Drugs 2022, 82, 251–274. [Google Scholar] [CrossRef]
- Gerace, E.; Landucci, E.; Bani, D.; Moroni, F.; Mannaioni, G.; Pellegrini-Giampietro, D.E. Glutamate Receptor-Mediated Neurotoxicity in a Model of Ethanol Dependence and Withdrawal in Rat Organotypic Hippocampal Slice Cultures. Front. Neurosci. 2018, 12, 1053. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Afanasyeva, M.A.; Van’kin, G.I. alpha-Synuclein knockout mice have cognitive impairments. Behav. Brain Res. 2012, 231, 226–230. [Google Scholar] [CrossRef]
- Mattson, S.N.; Crocker, N.; Nguyen, T.T. Fetal alcohol spectrum disorders: Neuropsychological and behavioral features. Neuropsychol. Rev. 2011, 21, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Olguin, S.L.; Thompson, S.M.; Young, J.W.; Brigman, J.L. Moderate prenatal alcohol exposure impairs cognitive control, but not attention, on a rodent touchscreen continuous performance task. Genes Brain Behav. 2021, 20, e12652. [Google Scholar] [CrossRef] [PubMed]
- Anokhin, P.K.; Shamakina, I.Y.; Ustyugov, A.A.; Bachurin, S.O.; Proskuryakova, T.V. A comparison of the expression of α-synuclein mRNA in the brain of rats with different levels of alcohol consumption. Neurochem. J. 2016, 10, 294–299. [Google Scholar] [CrossRef]
- Tehranian, R.; Montoya, S.E.; Van Laar, A.D.; Hastings, T.G.; Perez, R.G. Alpha-synuclein inhibits aromatic amino acid decarboxylase activity in dopaminergic cells. J. Neurochem. 2006, 99, 1188–1196. [Google Scholar] [CrossRef]
- Hausknecht, K.A.; Acheson, A.; Farrar, A.M.; Kieres, A.K.; Shen, R.Y.; Richards, J.B.; Sabol, K.E. Prenatal alcohol exposure causes attention deficits in male rats. Behav. Neurosci. 2005, 119, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, K.G.; Verma, P.; Yoon, E.; Yu, W.; Weinberg, J. Prenatal alcohol exposure increases vulnerability to stress and anxiety-like disorders in adulthood. Ann. N. Y. Acad. Sci. 2008, 1144, 154–175. [Google Scholar] [CrossRef]
- Chiavegatto, S.; Izidio, G.S.; Mendes-Lana, A.; Aneas, I.; Freitas, T.A.; Torrao, A.S.; Conceicao, I.M.; Britto, L.R.; Ramos, A. Expression of alpha-synuclein is increased in the hippocampus of rats with high levels of innate anxiety. Mol. Psychiatry 2009, 14, 894–905. [Google Scholar] [CrossRef]
- Pena-Oliver, Y.; Buchman, V.L.; Stephens, D.N. Lack of involvement of alpha-synuclein in unconditioned anxiety in mice. Behav. Brain Res. 2010, 209, 234–240. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Van’kin, G.I.; Bachurin, S.O.; Shamakina, I.Y. Differential involvement of the gamma-synuclein in cognitive abilities on the model of knockout mice. BMC Neurosci. 2013, 14, 53. [Google Scholar] [CrossRef]
- Chandra, S.; Fornai, F.; Kwon, H.B.; Yazdani, U.; Atasoy, D.; Liu, X.; Hammer, R.E.; Battaglia, G.; German, D.C.; Castillo, P.E.; et al. Double-knockout mice for alpha- and beta-synucleins: Effect on synaptic functions. Proc. Natl. Acad. Sci. USA 2004, 101, 14966–14971. [Google Scholar] [CrossRef]
- Connor-Robson, N.; Peters, O.M.; Millership, S.; Ninkina, N.; Buchman, V.L. Combinational losses of synucleins reveal their differential requirements for compensating age-dependent alterations in motor behavior and dopamine metabolism. Neurobiol. Aging 2016, 46, 107–112. [Google Scholar] [CrossRef]
- Carnazza, K.E.; Komer, L.E.; Xie, Y.X.; Pineda, A.; Briano, J.A.; Gao, V.; Na, Y.; Ramlall, T.; Buchman, V.L.; Eliezer, D.; et al. Synaptic vesicle binding of alpha-synuclein is modulated by beta- and gamma-synucleins. Cell Rep. 2022, 39, 110675. [Google Scholar] [CrossRef] [PubMed]
- Mak, S.K.; McCormack, A.L.; Langston, J.W.; Kordower, J.H.; Di Monte, D.A. Decreased alpha-synuclein expression in the aging mouse substantia nigra. Exp. Neurol. 2009, 220, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Pavia-Collado, R.; Rodriguez-Aller, R.; Alarcon-Aris, D.; Miquel-Rio, L.; Ruiz-Bronchal, E.; Paz, V.; Campa, L.; Galofre, M.; Sgambato, V.; Bortolozzi, A. Up and Down gamma-Synuclein Transcription in Dopamine Neurons Translates into Changes in Dopamine Neurotransmission and Behavioral Performance in Mice. Int. J. Mol. Sci. 2022, 23, 1807. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | |
|---|---|---|
| Forward | Reverse | |
| β-actin | 5′-cactgccg-catcctcttcct-3′ | 5′-aaccgctcatt-gccgatagtg-3′ |
| Snca (α-synuclein) | 5′-tgtcaagaaggaccagatg-3′ | 5′-caggctcatagtcttggtag-3′ |
| Sncb (β-synuclein) | 5′-agttccccacagacctgaag-3′ | 5′-ttacgcctctggctcgtattc-3′ |
| Sncg (γ-synuclein) | 5′-aaacatcgtggtcaccacc-3′ | 5′-tctagtctcctccactcttg-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokhan, V.S.; Chaprov, K.; Ninkina, N.N.; Anokhin, P.K.; Pakhlova, E.P.; Sarycheva, N.Y.; Shamakina, I.Y. Sex-Related Differences in Voluntary Alcohol Intake and mRNA Coding for Synucleins in the Brain of Adult Rats Prenatally Exposed to Alcohol. Biomedicines 2022, 10, 2163. https://doi.org/10.3390/biomedicines10092163
Kokhan VS, Chaprov K, Ninkina NN, Anokhin PK, Pakhlova EP, Sarycheva NY, Shamakina IY. Sex-Related Differences in Voluntary Alcohol Intake and mRNA Coding for Synucleins in the Brain of Adult Rats Prenatally Exposed to Alcohol. Biomedicines. 2022; 10(9):2163. https://doi.org/10.3390/biomedicines10092163
Chicago/Turabian StyleKokhan, Viktor S., Kirill Chaprov, Natalia N. Ninkina, Petr K. Anokhin, Ekaterina P. Pakhlova, Natalia Y. Sarycheva, and Inna Y. Shamakina. 2022. "Sex-Related Differences in Voluntary Alcohol Intake and mRNA Coding for Synucleins in the Brain of Adult Rats Prenatally Exposed to Alcohol" Biomedicines 10, no. 9: 2163. https://doi.org/10.3390/biomedicines10092163